COGNITIVE EVENT RELATED POTENTIALS DURING A LEARNING TASK
A thesis presented for the degree of
Doctor of Philosophy
Department of Clinical Neurological Sciences
Faculty of Medicine
University of Southampton
United Kingdom
By
Mohamed Fath EL-Bab
MB B Ch, MSc
April 2001
UNIVERSITY OF SOUTHAMPTON
ABSTRACT
FACULTY OF MEDICINE
CLINICAL NEUROLOGICAL SCIENCES
Doctor of Philosophy
COGNITIVE EVENT RELATED POTENTIALS DURING A LEARNING TASK
by MOHAMED FATH EL-BAB
Cognitive evoked potentials have been used to study a number of mental processes but little work has been on learning. This may be because the act of learning alters the nervous system and the experiments can not be repeated on the same subject.
This work examines the hypothesis that there are scalp recorded brain potentials in 99 healthy subjects learning a task.
Recording was by using cap electrodes with linked mastoids as reference. The volunteers was asked to observe a series of two hundred different images obeyed the rule for two patterns A and B, which had been generated by computer program and displayed randomly on a screen for 2sec and the screen was blank for 2sec. The images were classed according to the subject’s decision by pressing one of two buttons. Clues from border effect or contrast change were eliminated. One subject group was told the nature of the task beforehand some groups had feedback for correct/Incorrect answers and a control group simply observed the screen. Artefact due to eye movement, inattention and drowsiness were taken into account. Performance was monitored by the CUSUM quality control method. Learners were clearly distinct from non-learners. There were about half the subjects in each group.
Observers showed no significant electrical activity after the initial visual evoked potential, which ended at about 200msec. Learners showed a Positivity Associated with Learning (PAL) whether they learned or not. The PAL began at 200msec and lasted up to 2000msec. It was distributed over the frontal lobes but greater on the right side. Amplitude was about 6 m n and long lasting with no clear peak.
In general, the better the performance, the greater the positivity. Positive increased towards the end of a successful learning seen. Non-learners showed a similar positivity of lower amplitude. Positivity was greater after a single successful trial than after an incorrect answer. The frontal distribution of the PAL corresponds to metabolic studies by fMRI using a similar task. Our work gives temporal information and association between the potentials and subjects performance. It is proposed that similar studies will be useful in determining the pathophysiology of learning difficulties.
Dedicated to
All members of my beloved family
ACKNOWLEDGEMENTS
I would like to express sincere gratitude and deep thanks to all persons whose constant support and co-operation made the completion of the work possible.
I would like to thank my supervisor Professor Doctor E. M. Sedgwick for patience, kidness, encouragement, resignation at time, providing continuos helpful advice, and despite many horrendous personal problems.
I wish to thank Dr. Ian Philip for developing the computer software necessary for the experiment and continuos follow up for the equipments and recording
I would like to thank those volunteers who took part in the study for giving so much of their time.
I would like to express extreme gratitude and deep thanks to all the staff of the Department of Clinical Neurological Sciences in Particular Professor Fausto Iannoti, head of the department, and our secretaries Miss G. Cole and Mrs B. Mott, and all my colleagues for assistance, critical comments and inspiration.
I’m also indebted to my colleagues in the cognitive laboratory, in particular Professor Steven Harnad for the continuous expert advice, guidance, and being a very rich source of information in cognition.
I gratefully acknowledge the financial support of The Government of the Arab Republic of Egypt.
I’d like to express my deepest gratitude to my wife, Sherien and my sons Ahmed and Hesham for their practical and emotional support, patient, encouragement and Faith in my study and me.
Over all and finally unbounded thanks are to GOD for giving me everything which enabled me to establish this work.
Abbreviations
m
n = Micro VoltADC = analogue-to-digital converter
ANOVA = Analysis of Variance
ATL = left anterior temporal electrode
ATR = right anterior temporal electrode
Bit1 = trigger electrode one
Bit2 = trigger electrode two
C3 = Left central electrode
C4 = right central electrode
CA1 & CA3 = regions of Ammon’s horn in the hippocampus.
CI = Confidence intervals
CS = Conditioned Stimulus
CSD = Current Source Density
Cz = central midline electrode
DF = Degree of Freedom
df = degree of freedom
DT = Decision Time
EEG = Electroencephalogram
EPSP = excitatory post-synaptic potential
ERPs = Event related potentials
F = F ratio
F3 & F7 = Left frontal electrodes
F4 & F8 = Right frontal electrodes
FCZ = fronto-central midline electrode
Fig = Figure
FMRI = Functional Magnetic
Fr = with feedback and without rule
fr = without feedback and rule
FR = with feedback and Rule
fR = without feedback and with rule
Fz = frontal midline electrode
Heog = Horizontal electro-oculogram electrode
IPSP= inhibitory post-synaptic potential
K
W = KilohmL = Learner
Loc = Location
Lt = Left
LTD = long term depression
LTM = long term memory
LTP = long term potentiation
MRI = Magnetic Resonance Image
MS = mean square
msec = Millisecond
MTL = Medial Temporal lobe
MTL = medial temporal lobe
n = number of subjects
nL = Non-learner
NMDA = N-methyl-D-aspartate
O1 = left occipital electrode
O2 = right occipital electrode
Oz = Occipital midline electrode
P = P value
p value = probability value
P3 = left parietal electrode
P4 = right parietal electrode
PET = positron emission tomography
POZ = parito-occipital midline electrode
PZ= Parietal midline electrode
rCBF = regional cerebral blood flow
Resonance Image
Rt. = Right
SD = Standard deviation
SE = standard errors
Sec = second
SIG = significance
SS = sum of squares
STM = short term memory
T3 & T5 = left temporal electrodes
T4 & T6= right temporal electrodes
TPL = left temporoparietal electrode
TPR = right temporoparietal electrode
US = Unconditioned stimulus
Veog = Vertical electro-oculogram electrode
Aims and Objectives :
The human nature is to be curious about many mysterious things like, how we see, hear, move, learn, remember, and forget
a) Decision-making
b) Feedback according to whether the decision is correct or incorrect
Chapter 1: Introduction and Literature Review:
1.1. Historical aspect:
Diamond (1990) asked the leading Berkeley astronomer and Nobel Laureate Charles Townes "Which is more complex, the 100 billion stars in our galaxy or the 100 billion nerve cells in the 3-pound mass within our head?" Townes answered without hesitation, "the brain," "For, after all," Diamond responded "it is only the brain that can interpret our galaxy." And it is only the brain that can interpret its own cognizance.
Men ought to know that from nothing else but the brain come joys, delights, laughter, sports, sorrows, griefs, despondency, and lamentations. And by this, in an especial manner, we acquire wisdom and knowledge, see and hear know what are foul and what are fair, what are bad what are good, what are sweet and what are unsavory… And by the same organ we become made and delirious, and fears and terrors assail us… All these things we endure from the brain when it is not healthy…In these ways I am of the opinion that the brain exercises the greatest power in the man - Hippocrates, on the sacred disease (Fourth Century).
In 1848 in Berlin, Du Bois-Reymond recorded the standing potentials between the surface and the cut end of a nerve. The ‘action potential’ as we know now he described as a sudden negative variation in response to a stimulus.
Spontaneous EEG activity was first discovered in animal studies during the late nineteenth century in Liverpool by Caton (1875) using a Thomson reflecting galvanometer and Du Bois-Reymond’s coated, non-polarisable electrode. The Polish scientist Adolf Beck repeated many of Caton’s experiments and presented the electrical activity of the brain in 1890. He found that a spontaneous occipital oscillation disappeared with light stimulation, but not with noise. In 1929 Berger came up with a systematic description of the human EEG, but scientists at first had difficulty in accepting that his recordings were generated by, or even related to actual brain events. Berger speculated that the EEG was the manifestation of continuous psychological processes underlying non-localizable mental functions. It was not until Adrian and Mattews in 1934 validated his findings that the scientific community took Berger’s work seriously. From 1935 onward Gibbs et al established EEG as a clinical diagnostic tool in epilepsy.
Since then the technology has advanced and clinical applications such as epilepsy diagnosis has been developed. EEG activity has been assumed to represent underlying neural processes and the brain waves are generated by the synchronous electrical activity of literally millions potentials of individual neuron. By recording an EEG with certain stimuli or tasks it is tempting to speculate a cause and effect relationship. The actual process that scalp electrical potentials represent, is very difficult to establish due to the volume of unrelated and parallel activity that is occurring.
An evoked potential (EP) is the sequence of voltage changes generated in the brain, and in the sense organs and pathways leading to the brain, following the reception of the transient physical stimulus. It is represents electrical activity in those 100 billion nerve cells and is a useful research and clinical technique. Evoked potentials (EPs) developed from the electroencephalograph (EEG) that is used to record electrical brain activity. The EEG is a medical imaging technique that measures aspects of brain function by analyzing the scalp electrical activity generated by brain structures. It is a completely non-invasive procedure that can be applied repeatedly in patients, normal adults, and children with no risks or limitations. Both EP and EEG have the advantage of high temporal resolution but poor spatial resolution.
1.2. Normal Electroencephalogram:
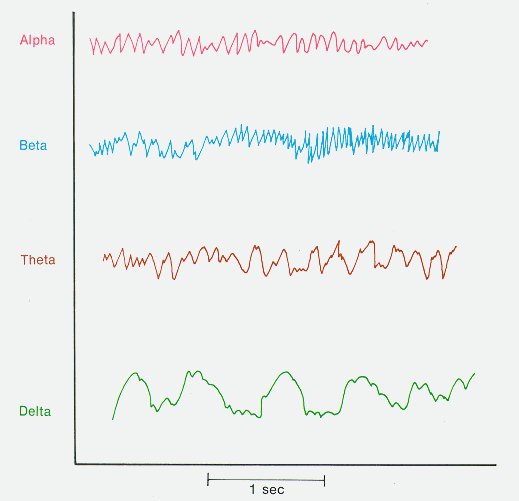
Four kinds of the brain waves are recognized from the scalp of normal individuals (Figure 1.1):
Figure 1.1 shows kinds of waves recorded in normal encephalogram (EEG)
Event related potentials (ERPs) studies are an attempt to eliminate unrelated potentials and noise. This is accomplished by time-locking the recording to the stimulus presentation and taking the average response to many stimuli. Only events that occur time and time again at the same point in the recording will actually show up in the final average. This assumes that little habituation occurs.
The evoked potentials are the electrical responses of the nervous system to motor or sensory stimulation with short latency potentials principally reflecting activity generated in the sensory receptors, a neural afferent pathway and its primary receiving area in the brain to a stimulus. These potentials are very small 0.5uv to 100uv- and they are usually recorded within the first 0.2 s of stimulus delivery. Sensitive amplifying equipment and the use of averaging technique (Dawson 1954) are required to detect these potentials, which are picked up at a considerable distance from the generator source, and have to be discriminated from other, usually much larger, potentials of physiological and environmental origin with which they are intermixed.
The term-evoked potential is defined as the average of multiple responses; the term-evoked response is defined as the electrical recording following a single stimulus. The term peak, or wave is defined as the positive (downward) or negative (upward) deflections from the baseline that make up the EP. The term component is defined as an individual contribution to the potential such as low-frequency component or late component.
1.3. The brain:
In order to understand how event related potentials illustrate brain functions, we should understand and know the brain cortical organization and structures (Figure 1.2).
The Egyptian papyruses were the first systematic written medical records, and the word brain first appeared in the Edwin Smith (1822-1906) translation of an Egyptian surgical papyrus. The brain basically consists of major parts; the brainstem, the cerebellum, the cerebrum, the diencephalon. All our conscious living such as thinking, memory, movement, consciousness, language, sensory perception, and emotion depend on the largest and complex part which is the cerebrum. It is divided into two hemispheres.
Two cerebral hemispheres have the same general appearance, and are incompletely separated by longitudinal cerebral fissure, at the bottom of this fissure they are united together by the corpus callosum. They are subdivided into four lobes frontal, parietal, temporal, and occipital. Each hemisphere has three surfaces lateral surface (convex), medial surface (flat), and inferior surface (very irregular). The gray matter (cortex), the white matter (axons), and the basal ganglia are the compartments of each hemisphere (figure 1.3a & b).
The right hemisphere is responsible for the simple language comprehension, the perception of spatial relationships, conceptual non-verbal ideas, general thought processes, and concentrates on the whole. The left hemisphere is active in speech, writing, calculation, language comprehension, analytical thought processes, and basically sorts out the parts. Carola et al. 1992 concluded that the left hemisphere sees the trees but not much of the forest, while the right hemisphere sees the forest but not too many of the trees.
Most people are right handed, and almost all of them have left-hemisphere dominance which generally speaking is more analytical, logical, precise and time-sensitive. While some left-handed people have right hemisphere dominance or mixed left and right dominance which is dreamier, more holistic and more involved with sensory perception and abstract cognition. The both sides of the brain are actively involved while anyone performing thought processes (Bryden, 1982).
1.4. Meninges of the brain (figure 1.4):
There are three connective tissue membranes covering the brain from outside to inside known by names as:
1.5. Cerebral Cortex:
The advanced intellectual functions of the human depend on the activity of the cerebral cortex and interaction of this structure with other portions of the nervous system.
The cerebral cortex was examined by the first microscopists, as early as 1776 and the first recorded structural detail was the stria in the occipital cortex, which was noted by Gennari, and named after him.
The cerebral cortex is involved in many aspects of memory, storage, and recall. It is very essential for musical and mathematical talents and for comprehension and execution of language. Higher cognitive function, and many complex motor activities depend on the cortex, and it is responsible for the perception and conscious understanding of all sensations, and it is a site in which any modality of sensation can be integrated with others.
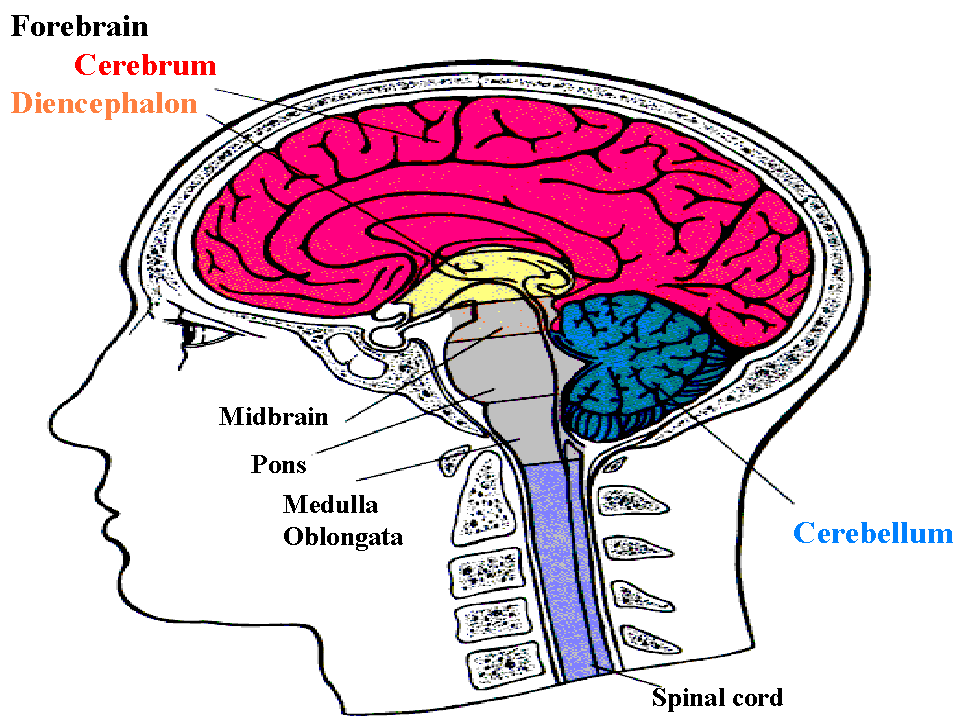
Figure (1.2) shows side view of the human brain location in relation to the skull, the spinal column and the spinal cord.
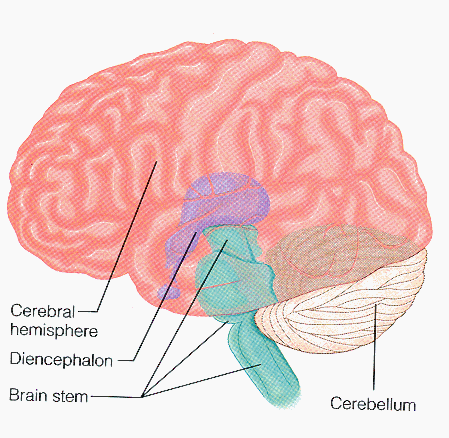
(a) (b)
Figure (1.3) shows the brain compartments and relations (a) left lateral view and (b) superior view
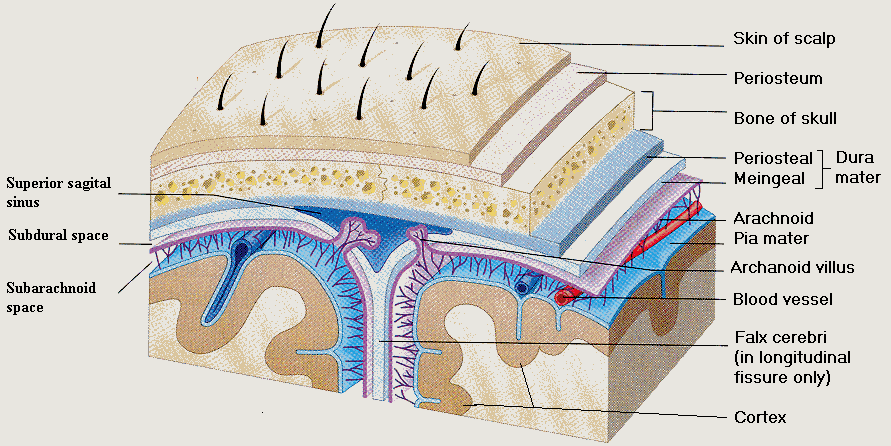
Figure (1.4) shows the brain covers starting from outside the Skin, the Skull, and the Meningese.
1.6. The Cerebral Cortex Structures (Figure 1.5):
To the unaided eye it forms a complete mantle ‘pallium’ covering the hemisphere and obviously variable in the thickness (1.5 to 4.5 mm). It is thicker on the exposed convexities of gyri than in the depths of sulci. The cerebral cortex containing over 100 billion neurons and 250 billion glial cells.
The cerebral cortex can be divided into five types based on its structure complexity. By the largest part is the neocortex. Whichever piece of cortex is examined, it is all constructed on the same plan. There are six recognizable areas arranged in laminated manner, superficial to deep (Brodmann 1909) as follows:-
Careful examination of the thickness and number of cells in these layers reveal systematic differenceson the basis of these differences brodmann 1909 recognised 52 areas. Many of these areas have specific functions; areas 1, 2 & 3 are primary sensory cortex, area 17 is visual and area 4, the motor cortex.
As well as the organisation in layers, there is an organisation in radial columns. Afferent and efferent fibers run radially, neurons in a radial column tend to respond to different aspects of the name thing e.g. movement in one direction.
The afferent fibers to the cortex run radially towards the surface and synapse in Layer one through layer four. Projection fibers come from the Thalamus, association fibers from widely dispersed areas, and the third group from several specific subcortical structures outside the Thalamus. These include the locus ceruleus (origin of noradrenergic fibers), the raphe nuclei of the brain stem (origin of serotonergic projections), and the basal nucleus of Meynert in the basal telencephalon (origin of cholinergic projections). The corpus callosum and anterior commissure contain commissural fibers that link corresponding and non-corresponding regions of the two hemispheres. The efferensts projecting to the brain stem and spinal cord arise from layers V and VI.
The neurons of the pallium have been described and categorised into different classes; the great majority falling into these cell types.
Pyramidal cells named from the shape of their somata, varying from small elements measuring about 10m m across to the giant pyramidal cells (of Betz) measuring up to 70m m or more. The pyramidal cells in layer II, III, and V, serve the major efferent pathway of the cerebral cortex. Their apices are oriented towards the surface of the cortex. The small pyramidal cells in layers II and III project to other cortical regions. The projection axons of the large pyramidal cells in layer V extend centripetally out of the cortex to reach more or less distant subcortical structures, such as basal nuclei, brainstem nuclei, and the grey matter of the spinal cord. The so-called pleomorphic cells are considered as modified pyramidal cells with axon entering the white matter. Their somata are variably shaped. The pyramidal cells are perpendicular to the cortical surface with dendrites long enough to form effective dipoles. The perpendicular spatial organization leads to the summation of the associated currents, especially when these neurons are synchronously activated. Non-synchronous activity and activity from radially symmetrical stellate cells do not appear in EEG or ERPs. Most of the evidence available at present suggests that the scalp recorded potentials are due to excitatory (depolarising) or inhibitory (hyperpolarising) postsynaptic potentials developed by the cell body and dendrites of pyramidal neurons, rather than axonal action potentials (Allison et al. 1986).
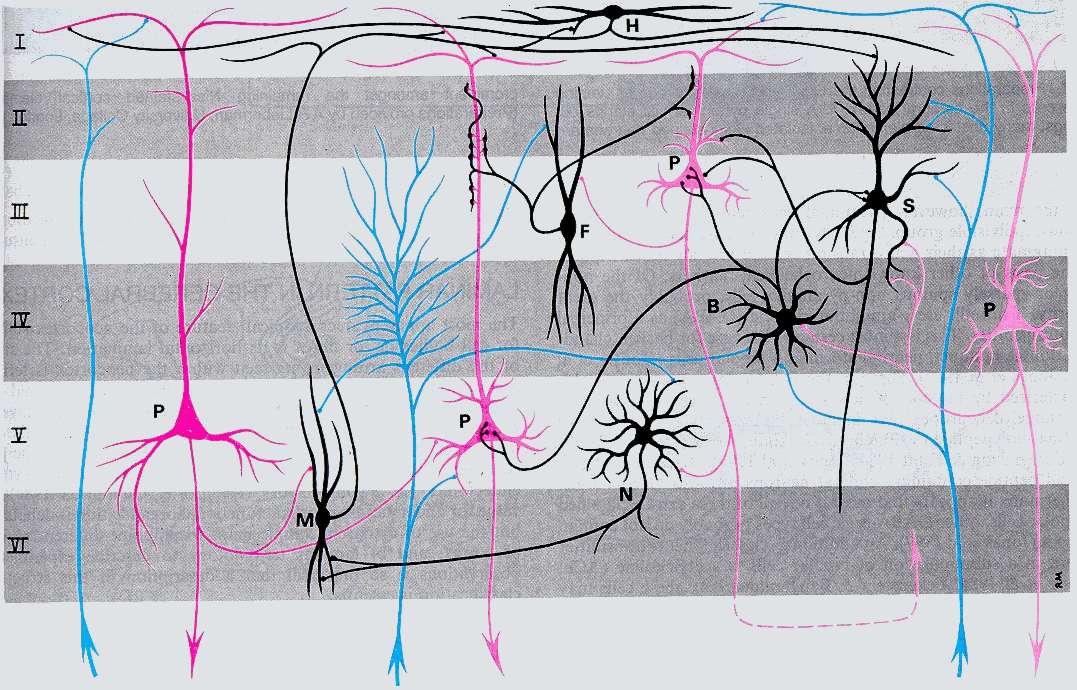
Figure 1.5. Cerebral cortex layers and the most frequent types of neo-cortical neurons, connections with each others and afferent fibers (blue). Neurons (black). Efferent (Magenta), Pyramidal (P), Fusiform (F), Horizontal (H), Neurogliaform (N), Martinotti (M), Basket (B), Stellate (S)
1.7. Physiological basis:
Neurons collect, process, and relay information by generating electrical signals that are transmitted via the cell axon to synaptic junctions with other cells. Their ability to perform this complex task is due to the unequal distribution of electrically charged particles on either side of its semi-permeable cell membrane.
A neuron forms synaptic connections with axon terminals from many different nerve cells. Several excitatory as well as inhibitory ionic disturbances can therefore occur simultaneously, or nearly so, in neighboring portions of the post-synaptic cell membrane. Both temporal and spatial summation of excitatory and inhibitory post-synaptic potentials occurs. The outcome of these interactions determines the size of the membrane potential at any point in time.
It is often said that humans consist of 2/3 water. Strictly speaking, that should be 2/3 salt water, the principal salts being the positively charged cations of sodium (Na+); potassium (K+), calcium (Ca2+) and magnesium (Mg+) and the negatively charged anions of chloride (Cl-) and phosphate (P-) and various organic acids (A-). All of these chemicals (and more) are contained in both the extracellular fluid surrounding each neuron, and intercellular fluids (cytosol) of neurons, and both contain high concentrations of compounds, called electrolytes, that in solution, conduct an electric current and are decomposed by it into atoms called ions capable of carrying positive or negative electrical charges. Their concentrations are not the same. The fluid inside neurons contains more negative anions and/or fewer positive cations than the surrounding extracellular fluid from which it is separated by the semipermeable cell membrane. (The permeability of this membrane to specific ions is determined by the presence of specific ionic channels.) The unequal distribution of positive and negative charges between the inside and outside a neuron results in an electrical potential of -70 mV. This resting membrane potential of the cell comes about because of interplay of several forces including (1) diffusion, (2) electrostatic pressure; and (3) active sodium and calcium pumps.
The preceding discussion describes the neuron at rest. To do its job of collecting, processing and distributing information, the cell's membrane potential must be disturbed. In humans, and other mammals, this is most commonly instigated or "stimulated" by the release of a chemical transmitter substance from an axon terminal. The neurotransmitter diffuses across the synaptic cleft and acts on chemical receptor s on the membrane of a dendrite or the cell body of the post-synaptic neuron. The action of the transmitter chemical on the special protein receptor structures of the post-synaptic membrane opens the gates of "transmitter-dependent" ion channels. This permits a particular type of ion to pass through the cell membrane, thus changing the local membrane potential
There are two modes of transmitter action. In the simpler case, the neurotransmitter acts directly on the gates of ion channels to allow the influx or efflux of a particular ion (e.g., sodium). A more complex process involves the activation of G-proteins that may open ionic gates directly or stimulate the synthesis of a second messenger chemical (the neurotransmitter is the first) in the cytoplasm of the cell. The second messenger then starts a cascade of chemical reactions to open specific ion channels. The action of second messengers, like that of some neurotransmitters, is usually terminated by rapid enzymatic destruction of the second messenger. In many neurons, the second messenger is cyclic adenosine monophosphate (cyclic AMP) which is rapidly destroyed by the enzyme phosphodiesterase.
These transmitter/receptor interactions may increase or decrease the potential difference between the inside and the outside of a cell, depending on the type of ion channel that is affected. An increase in the influx of positively charged cations, such as sodium or calcium, reduces the membrane potential and thus depolarizes a portion of the post-synaptic membrane. This results in an excitatory post-synaptic potential (EPSP). An influx of negatively charged anions, such as chloride or an efflux of positively charged cations, such as potassium, increases the local membrane potential and thus hyperpolarizes the postsynaptic membrane. This produces an inhibitory post-synaptic potential (IPSP).
Both EPSPs and IPSPs are low-amplitude electrotonic potentials (tonic = tension) that are propagated passively and hence decrementally (their size decreases) along the neuronal cell membrane. They are "graded" potentials - i.e., their amplitude reflects the intensity and duration of the interaction between the neurotransmitter and its receptor complex. Neighboring EPSPs and IPSPs interact (i.e., sum and subtract).
Both EPSPs and IPSPs are very transient phenomena, their duration being determined by the length of the transmitter/receptor interaction or the life span of the second messenger. Neurotransmitters are quickly removed from the synaptic cleft, most commonly by re-uptake into the axon terminal that secreted them, although enzymatic destruction of the transmitter molecule terminates the action of some neurotransmitters. The most common second messenger, cyclic AMP, is also very rapidly destroyed enzymatically. Some second messengers have more persistent effects, thus prolonging the effective action of some neurotransmitters.
1.9. Electro-physiological basis:
The potentials of the brain recorded in an EEG appear in waves ranging from 1 to 40 or more cycles per second (hertz or Hz), with amplitude ranging from -100 to +100 microvolts. The amplitude of the EEG signal strongly depends on how synchronous the activity of the underlying neurons is.
Each activity is the result of electrical and physiochemical changes in the brain tissue. The external world is transmitted to the cortex via sense organs and the characteristics of these organs and the transmission pathways are reasonably well understood. However, what happens once the sensory activity reaches the cortex is still poorly understood. The transmission of information to the brain involves the flow of ions across the neuronal membrane producing a voltage field around each active neuron. The potential difference between the postsynaptic membrane portion and the other parts of the neuronal membrane causes an electrical current to flow along the cell body and dendrites with a return current in the extracellular space. These electrical potentials summate in the cortex and extend through the coverings of the brain to the scalp. These spontaneous electrical activities are known as electroencephalogram (EEG) and by using the electroencephalographic technique we are able to record these voltage changes.
Although one neuron generates too small a signal to be observed at any considerable distance from the nerve cell, hundreds of thousands of them can produce a potential field that is strong enough to be detected at the scalp. When a group of cells is excited simultaneously, the tiny signals combine to generate one large surface signal. The electric conductivity of the skull, which separates the brain from the scalp electrodes, is low enough for an electric current to pass through (Figure 1.6)

Figure (1.6) shows the generation of very small electrical fields by synaptic currents in pyramidal cells. The EEG electrode measures this signal through the thick layers of tissue. Only if thousand of cells simultaneously their small voltage can the signals become large enough to be seen at the surface of the scalp.
1.10. Event related potentials:
1.10.1. Event related potentials (ERPs)
Event related potentials (ERPs) recorded from scalp electrodes have been used widely to study human cognitive processes and their neural substrate.
Event-related potentials (ERPs) are small voltage fluctuations resulting from evoked neural activity. These electrical changes are extracted from scalp recordings by computer averaging epochs (recording periods) of EEG time-locked to repeated occurrences of sensory, cognitive, or motor events. The spontaneous background EEG fluctuations, which are random relative to the stimuli, are averaged out, leaving the event-related brain potentials. These electrical signals reflect only that activity which is consistently associated with the stimulus processing in a time-locked way.
We assume that the EP (signal) has known time relationship to the stimulus whereas background brain activity (noise) does not. Averaging is the presentation of a stimulus many times and signals for the duration of interest immediately following are summed before being divided by the number of presentations to give the average EP. The aim is to improve the signal to noise ratio (S/N ratio) so that the EP is more discernible. S/N ratio improves by factor of the square root of the number of presentations (Table 1.1); (Regan 1989)
|
Number of presentation |
Signal ( m n ) |
Noise ( m n ) |
S/N ratio |
|
1 |
1 |
1 |
1:1 |
|
4 |
1 |
½ |
2:1 |
|
9 |
1 |
1/3 |
3:1 |
|
16 |
1 |
¼ |
4:1 |
|
81 |
1 |
1/9 |
9:1 |
Table (1.1) shows the signal to noise ratio (S/N ratio) improvements.
We only need 9 presentations to treble the S/N ratio but a further 72 (i.e. 81 total) presentations are needed to triple the ratio again.
The nomenclature of EP entails labeling by their polarity and latency. "N" and ‘P’ refer to negative and positive polarity respectively. A number denoting the latency in milliseconds follows this.
Clinically, the most useful response is obtained following, stimulation of the sensory modalities for vision audition (Auditory Evoked Potentials. AEP) (Figure 1.7), (Visual Evoked Potentials. VEP) (Figure 1.8), and bodily sensation (Somatosensory Evoked Potentials. SEP) (Figure 1.9). These early, short latency potentials principally reflect activity generated in the afferent pathway and its primary receiving area in the brain. These potentials vary as the physical characteristics of the stimulus vary. There are also potentials, occurring at longer latency, which appear to influenced by mental processes such as attention to the stimulus or
expectation that particular stimulus will occur. These are the potentials, which I am interested in and these are termed "endogenous or event related potentials" (ERPs) to distinguish them from the evoked potentials elicited by external physical events (Halliday 1992).It is possible also record a longer latency ERP to a non-event, such as the omission of an expected stimulus from regular train. Such ERPs have been termed ‘emitted potentials’, since there is no outer event to the trigger them (Picton, 1988).
Other ERPs may be recorded in association with awaiting an expected stimulus (‘expectancy wave’ or ‘contingent negative variation’, CNV) or when preparing to make a motor response (‘readiness potential’); these are recorded as gradually rising negativities located over the central cortex for 1 or 2s before the relevant event.
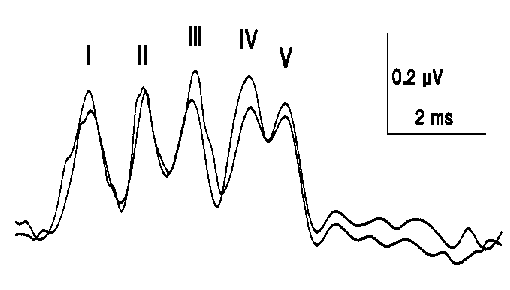
Figure (1.7) Normal Brainstem Auditory Evoked Potential to stimulation of the right ear, recorded between vertex (CZ), and right ear (A2) top tracing. Bottom tracing between vertex and left ear (A1).
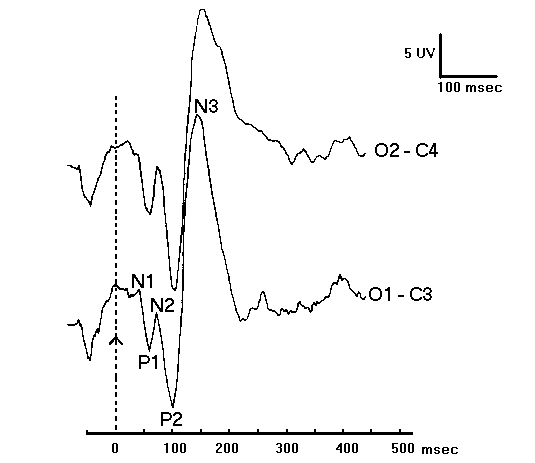
Figure (1.8) shows the normal Visual Evoked Potential (VEP), to diffuse light flash. The VEP divided into a primary response and secondary response. Negativity at the occipital plotted upward.
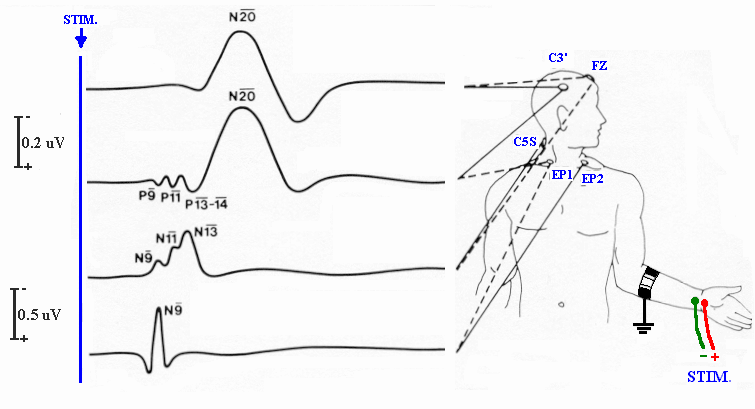
Figure (1.9) shows the normal Somatosensory Evoked Potential (SEP) to arm stimulation, from top to bottom, Far field recorded between scalp and cephalic reference electrodes, and between scalp and non-cephalic reference electrode. (N20), Erb’s point potential N9, and Cervical SEP N13.
1.10.2. Early event related potentials:
The first waveforms in the visual are the N100 and P100, which represent the physiological action of the visual system. The P100 is thought to represent visual cortical activity.
The earliest components of the EP are far field potentials and they reflect activity in the receptors and peripheral relay stations. (Calloway et al 1975). Following these are the early cortical evoked responses, which appear to be generated in the primary receiving areas of the brain. Example include N100 and P100 (both occurring 100msec after presentation of a visual stimulus) which reflect activity of the visual pathways and cortex. Far field and early cortical evoked potentials only reflect the physical properties of the stimulus and are insensitive to the succeeding cognitive processes.
First of three components in visual discrimination tasks is a posterior N200 wave and has been named ‘selection negativity’ (Czigler, 1995). It has been shown to increase in amplitude to task deviant stimuli. Secondly an anterior positivity to task related stimuli occurs around 200 msec its behavior is to increase to correctly recognized stimuli. Both the selection negativity and the P200 are dependent on the on-line processes that identify relevant visual features. The third component is the N400, which thought to be correlate of stimulus categorization or orientation and is increased by task relevant deviant stimuli. (Czigler, 1995).
The evoked potentials that are determined by physical aspects of the stimulus have been labeled, ‘exogenous’, appearing in the first 80 msec of recording. Up to 200 msec after a stimulus sensory-encoding is occurring. After 200 msec the cognitive components of the ERP are observed. Labeling the components is by latency and polarity. A positive component at 100 msec is called ‘P100’ and a negative deflection at 200 msec is a ‘N200’. Many of the cognitive waves are named after their latency when first seen, these names have stuck, e.g. a ‘P300’ can occur at any point between 300-800 msec and consequently the ‘N400’ may occur prior to it. (Figure 1.10) of course the electrodes from which a potential is recorded must also be known.
In contrast, Event related potentials (ERP) which occur after 200msec are of considerable scientific interest in this study because of their relationship to cognitive functioning. The ERP waveform is normally viewed as a series of components, each of which is a manifestation of the synchronized activity of a population of neurons (Ducan-Johnson and Dochin 1982). In general, scientists believe that learning involves an alteration in the functional properties of neurons and synapses (Ciesielski and French 1992). ERP could supply a long sought after physiological correlate of learning but as of yet there has been very little research.
1.10.3. Late event related potentials:
It is the study of the ERP components that are related to higher cortical functions, and thus have captured the interest of researchers interested in perceptual, cognitive, and motor behavior.
1965 Sutton et al first described P300 and related potentials associated with cognitive function.
The known parameters governing the form of P300 are
The P300 wave is a positive peak with latency of approximately 250-800 msec of the human event related potential (ERP) has been typically characterized in terms of latency, amplitude and scalp amplitude distribution. P3 latency has been shown to covary with speed of information processing as indexed by reaction times, the faster speed of processing, the earlier P3 latency (Ritter et al. 1972; Picton et al. 1974; Ford et al. 1976; Courchesne et al; 1977 & Kutase and Donchin, 1978).
The latency of P300 increases with the time the subject needs to distinguish the rare stimulus. The amplitude increases with rarity of the stimulus and to some extent with stimulus intensity. This is a late wave positivity that occurs with a variety of stimuli in many experimental conditions. It has been linked with many processes and has been classically evoked with the oddball paradigm. This is where two stimuli are presented one of which is an infrequent attended target. When the target is attentionally discriminated from the other stimuli, by being counted for example, the P300 is of greater amplitude (Yamaguchi and Knight, 1995; Polich, 1989 & 1990; Duncan- Johnson and Donchin, 1977). In normal young adults a positive wave over the Centro-parietal scalp is seen. A P300 can be provoked in multi-stimuli paradigms.
As well as the attention dependent P300 a passive or ‘novelty’ P300 occurs to infrequent non-target stimuli (Courchesne et al, 1984; Knight, 1984). The characteristics of the wave are different in that it is of shorter latency (250msec), of frontal-central recording sites and habituates rapidly (Polish, 1994). This waveform has been titled the P300a to distinguish it from P300 (now P300b). Most data exists in the form of auditory trials. In different modalities it as been reported as Centro-parietal (Pfefferbaum et al, 1980 & 1984), and also a later parietal potential (Courchesne, 1978).
The amplitude of the P300 has been shown to be inversely proportional to stimulus presentation probability and directly to task complexity (Johnson, 1988). Visually patients with temporoparietal lesions show a decreased novelty P300 with no target decrement (Yamaguchi and Knight, 1995). Different generators of cortical circuits are engaged in target or novelty P300 generation.
P3 amplitude has been shown to be inversely related to a prior probability, the lower probability, and the higher the P3 amplitude. (Tueting et al. 1971; Squire et al. 1973,1975a; Ford et al. 1976b; Picton et al. 1976; Duncan-Johnson and Donchin, 1977), and to be influenced by the sequence of immediately preceding events, i.e., sequential event structure (Squire et al. 1976; Duncan- Johnson and Donchin, 1977).
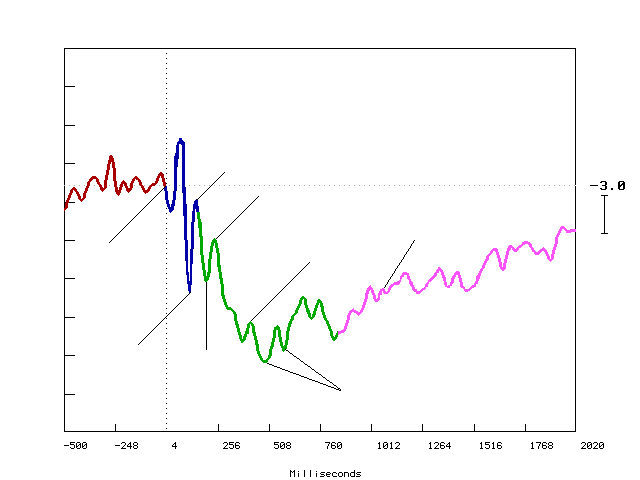
Figure (1.10) shows an example of the Event Related Potential ERP recorded from mid-frontal site at FZ electrode during a learning task. Y-axis shows amplitude in Microvolts (
m n ), and X-axis shows latency in milliseconds (ms). The black vertical dotted line marks the point of stimulus onset, and the horizontal black dotted line represents the baseline. Downward deflection represents the positivity. Upward deflection represents the negativity.The P300 has a wide distribution with maximum in parietotemporal areas, unrelated to the specific sensory areas but probably related to the parietotemporal association cortex and subcortical structures such as hippocampus and thalamus. The P300 to novel stimulus may be located more frontally.
Differences in P3 scalp amplitude distribution have been related to differences in the cognitive processes underlying the generation of these waves (Courchesne et al. 1975; squire et al. 1975b; Stuss and Picton 1978).
Across time P3 scalp distributions do not change so long as the task and eliciting event category do not change, and the effect of the prior probability and sequential event structure on P3 amplitude are relatively constant across time. That is the subject has been considered to be passive and a consistent responder to the event categories, the event probabilities and the task specified by the experimenter. These views of P3 waves and their underlying cognitive processes do not take into account the dynamic nature of human information processing, a given event may later generate notions about past as well as future events. When such alterations occur, the cognitive processes is brought to bear on succeeding events which might differ from those brought to bear on past (earlier) events.
If different P3 scalp amplitude distributions do reflect different cognitive processes, then it is conceivable that changes in P3 amplitude and distribution might be found when subjective notions about an event category are altered. Some researchers (Courchesne et al. 1975, 1977 &1978. Courchesne 1977) have suggested that very novel events elicit frontal P3 waves while less novel events elicit parietal waves. The subjects expressed surprise and interest over initial presentations of both very novel and less novel events, but indicated that they soon began to expect them and, finally to ignore them. These findings suggest that (1) successive presentation of novel events should elicit successively less frontally and more parietally maximal P3 waves, and (2) both novel and less novel events should elicit successively smaller P3 waves as subjects come to ignore them. These two possibilities are inconsistent with the two views current in literature, which were mentioned above.
Mecklinger and Ullsperger (1995) hypothesized that the relationship between P300 amplitude and categorization tasks might be modulated by ease of assignment to category. e.g. after learning the task the P300 increased. When categories are distinct enough to easily classify stimuli the P300 amplitude reflects the chance of target category. In other words before an internal category representation is formed the P300 reflects the ease of stimulus category designation. Whereas after learning the target probability is the deciding factor. These attributes of P300 will be of importance in our experiments.
The P300 name is somewhat misleading since it may occur anywhere between 300msec and 1000msec (Rosler et al 1986). It is a composite potential which appears to have numerous component entities e.g. P3a, P3b, and slow wave. Learning may be linked to changes in ERP and it has been reported that learning evokes specific changes within the domain of the P300 (Rosler et al 1981).
Amplitude and latency of ERP components can be used as indices of the nature and timing of a subject’s cognitive response to stimulus. The two parameters of P300- latency and amplitude are believed to provide indices of information processing (Johnson et al 1985). P300 amplitude is a measure of the amount of information processing. It reflects the amount of information carried by the stimulus (Looren et al 1988 and Woestenburg et al 1992). P300 supplies information about the chronometry of information processing (Kramer and Strayer 1988). P300 latency is proportional to the time it takes the subject to evaluate and categorize the stimulus (Horst et al 1980 and Ducan-Johnson 1981).
P300 latency is age dependent, being longer in children, progressively decreasing until 18 before increasing by 1.25msec per year (Barajas 1991 and Diniz and Fukuda 1997).
In adult, aging increases the latency, decrease the amplitude and cause forward shift in the distribution of P300 (Misulis 1994)
A large number of clinical studies of the P3 component have demonstrated reduction in P3 amplitude in individuals expressing a wide range of behavioral dysfunction, one of which is memory impairment (Rugg, et al. 1990).
The ERP thus reflects, with high temporal resolution, the patterns of neuronal activity evoked by a stimulus. Due to their high temporal resolution, ERPs provide unique and important timing information about brain processing. Mental operations, such as those involved in perception, selective attention, language processing, and memory, proceed over time ranges in the order of tens or hundreds of milliseconds. Most other functional imaging techniques require the integrating evoked brain activity over many seconds and are thus unable to capture the time sequence of these operations. ERP recordings, however, provide a millisecond-by-millisecond reflection of evoked brain activity.
For this reason, ERPs are an ideal methodology for studying the timing aspects of both normal and abnormal cognitive processes.
On the other hand, ERP data provide less accurate spatial information than positron emission tomography (PET) or functional magnetic resonance imaging (fMRI), which lack fine temporal resolution. As a result, ERPs represent the natural complement of PET and fMRI to study human cognition. Whereas PET and fMRI can localize regions of activation during a given mental task, event related potentials ERPs can help in defining the time course of these activations.
PET studies provide a consistent set of data for the localization of areas active during learning, although they cannot provide the extraordinary time resolution.
Mapping human cognitive function is a major emphasis of ERP studies at our department. Research will help explore the organization of cognitive processes such as selective attention, memory, language, and learning both in normal In a clinical setting, the temporal resolution of ERPs is useful in identifying at which level along the sensory pathways a lesion is localized. Visual ERPs help the early diagnosis of multiple sclerosis before any structural abnormality is detected. ERPs are also used to monitor comatose patients to evaluate the functionality of vital centers in the brainstem.
1.10.4. Learning & Memory and the P300
Learning probably involves many different parts of the brain and no one has been able to say with any degree of certainty where it occurs (Hilgard et al 1966). Learning establishes long-term memory during learning (Sqiure and Zol-morgan 1991), and the process involves many brain structures including the frontal lobe.
In a study of ERPs evoked by the Von Restorff effect on memory recall, Karis et al. (1984) established striking differences between subject's strategies they used to perform the memory task. The Von Restorff effect can be described briefly as an increased recall probability for an item which different from others in an obvious way (e.g. in size, color, class or nature) in a list of items to be remembered (this different items called an isolate). Fabiani et al. (1990) confirmed the observation of Karis et al. (1984). These differences make a novel task relevant target. Earlier data suggested that the P300 reflected processes invoked when there is a need for context updating i.e. evaluation of current representation in working memory (Donchin, 1981; Nageishi and Shimokochi, 1980). This is not far from the orienting response (Polish, 1988; Donchin, 1986).
The effect is larger with decreased probabilities and tempered by time. The interval between stimuli suggests P300 is sensitive to the strength of a decaying memory representation. At short inter stimuli intervals only rare targets need updating upon representation as frequent event are likely to occur while their previous image is still held in working memory. The amplitude is related to the overall degree of updating that is required and may be a global closure of encoding (Fiztgerald and Picton, 1981). Donchin (1981) stipulated that P300 is a manifestation of processes that maintain an accurate environmental model, or schema, by continually revising this modal according to most recent useful sensory input.
P300 amplitude is proportional to the amount of attentional resource allocated when the stimuli are processed (Polish et al, 1994; Siddle, 1991). The latency is associated with the speed of stimuli to class during memory update (Duncan-Johnson, 1981; Magliero, 1984).
In agreement with Von Restorff effect, memory for events that elicit a large P300 is better than otherwise (Karis et al, 1984 and Fabiani et al, 1986). Consequently larger amplitude is measured in a memory test to repeat items and to correctly identify old items. Older studies also show this increase with the degree of confidence with which the decision is made (Squire et al, 1975a and 1975b; Paul and Sutton, 1972).
Generation of the P300 is dependent on an inter-hemispheric interaction in the temporal parietal junction (Yamaguchi and Knight, 1995). This leads to the conclusion that the P300 response is dependent on association cortex with the limbic system, sustaining and updating a model of external envelopment (Donchin, 1979). Could the novelty P300 be a momentary shift of attention towards an unexpected event in the working memory trace (Ford et al, 1976c; Näätänen and Gaillard, 1983)? The generation has been suggested to be a summation of similarly active neural generators each faithful to a specific cognitive processes.
1.10.5. The medial temporal lobe and the P300:
The medial temporal lobe (MTL) consists of the hippocampus, amygdala and parahippocampal gyrus and has been linked to recent event memory. Bilateral damage to this area causes permanent anterograde global amnesia (Scoville and Milner, 1957; Squire, 1986). Brief disruption of the MTL during the first 650msec following stimulus presentation for encoding leads to decreased performance in recall tasks (Halgren et al, 1985). This shows the time course for the MTL encoding, integration and storage contribution to memory.
The P300, which actually appears around 600msec increases with stimulus repetition and thought to reflect closure of the completed encoding/integration, process (Altafullah et al, 1986). In these terms the closure of encoding or contextual integration is active (Heit et al, 1990).
A MTL P300a occurs to infrequent unattended stimuli but is too early to be mistaken for the P300b. Performance in an oddball paradigm is not altered by unilateral anterior temporal lobectomy (Stapleton et al, 1987). MTL activity tasks require recent contextual memory, no lexical decision nor sensory discrimination effect can be seen.
1.10.6. N400:
Numerous studies that have employed a task in which items were presented sequentially and subjects were asked to respond if the stimulus was unmatched (incongruent) or matched (congruent) with the preceding items (Barrett, and Rugg, 1989, 1990). A late negative component, peaking around 400 msec, in frontal area is observed and is larger in non-matched items than matched items. It has been proposed that it is represent the associated activation of neural networks basic to stimulus integration (Halgren and Smith, 1987). The N400 is larger to non-repeated words or faces and semantically incongruent words at the end of a sentence (Kutas and Hillyard, 1980, Iragui et al, 1996). The P300 on the other hand is maximal to repeated words and attended infrequent tones (Smith and Stapleton, 1986; Halgren and Smith, 1987).
It is interesting to know that N400 component, as recorded from the scalp is severely attenuated following unilateral anterior temporal lobectomy (Smith and Halgren, 1988) suggesting that the temporal lobes may have an important role in eliciting N400.
The N400 has been hypothesized to represent a construction of a cognitive gestalt that encodes the stimulus and its prior contextual occurrences through associative activation of a neocortical MTL network (Halgren and Smith, 1987). Some evidence for this is seen in the decreased amplitude and duration to repeated stimuli (Smith et al, 1986). This repetition sensitivity is lost after left anterior temporal lobectomy (Smith and Halgren, 1989).
1.10.7. Abnormal P300
It is well known that bilateral lesions of the human medial temporal lobes produce severe amnesia for recent events (Scoville and Milner, 1957; Squire, 1982). Causes of amnesia syndromes include thiamin deficiency, resulting from alcohol abuse or severe malnutrition or malabsorption, head injury post-herpes encephalitis all of that can cause severe damage to the medial aspect of the temporal lobes as well as other damage. Severe hypoxia, deep midline tumors and posterior cerebral artery occlusion, Alzheimer’s disease, etc. can all cause amnesia.
It has been reported P300 latency to be prolonged in adult patients with mental retardation, in patients with dementia of various causes, in patients with parkinson’s disease, in chronic schizophrenic patients.
Patient with temporal lobe epilepsy may have delayed P300 which is independent of seizure manifestation or antiepleptic drugs (Fukai 1990).
Abnormalities in P300 latency may develop in early Alzheimer’s disease and progress proportionately to intellectual decline (Ball et al 1989, and Polich et al 1990)
Delayed P300 has been reported in patients with AIDS and AIDS-related complex even before psychometric tests could detect cognitive deficits (Ollo et al 1991). P300 amplitude changes have been described in various conditions, including frontal lobe lesion, hyperactive children treated with methylphenidate, increase blood lead levels in children, infantile autism, and schizophrenia.
1.11. Learning
1.11.1. Definitions:
The world of science, like that art or religion, is a world created by the human imagination, but within very strict constraints imposed both by nature and the human brain (Jacob. 1988).
What exactly is the definition of learning and memory? Learning and memory are different sides of the same coin. Memories are what left behind as a result of learning, and we infer the existence of learning from the presence of memories. What exactly do we mean by the term of learning and memory? The definition of these apparently innocuous terms has been a topic of passionate debate by psychologists.
The definitions are numerous and few satisfactory for this multifactorial process but perhaps the most comprehensive I have encountered is:
Learning is the storage of the information as a function of experience and resulting in a relatively permanent change in personality (including cognitive, affective, attitudinal, behavioral, experiential, and the like) and reflect a changes in performance usually brought about by practice although it may arise from insight or other factors including memory. Which we mean any process whereby a person or a machine increases its knowledge or improves its skill.
This is a good reflection of the multitude of components involved in learning and is not within the scope of this treatise to comment on all of them. Rather, a synopsis of what are believed to be the fundamental features will be presented.
Memory is learning‘s faithful partner. It refers to stored information produced by learning, and is an abstract term that describes mental states which carry information, while learning describes a transition from our mental state to a second, in which the information is in some way different.
When we say that the dog in a laboratory Pavlovian conditioning experiment has learned and remembers something about the relation between food and bell, what we mean is just that a new behavior has been conditioned: the dog salivates to the bell, whereas previously it did not. On such a view, we should only consider the term of ‘leaning’ and ‘memory’ if there is some observable change in behavior, in which case the new behavior is the learning and memory
However, there are two obvious problems with that definition. The first is that learning may occur without any concomitant change in behavior; if a CS and US such as a shock are presented to subject administered with drugs that block muscular activity, conditioned responses may perfectly well occur to the conditioned stimulus when the paralytic drug has worn off (Solomon and Turner, 1962).
Learning clearly occurs when the animals are paralyzed, even though no behavioral changes take place at that time. The second problem is that many in cases it can be established that organisms do much more than simply acquire new types of behavior. For instant, in a famous experiment, MacFarlane (1930) trained laboratory rats to run through a maze to obtain food, and found that when the maze was filled with water, the animals continued to take the right path to the food even though they now had to swim to reach it. Clearly, learning in this case does not merely involve equitation of a set of particular muscle activities conditioned to a set of stimuli, but instead involves acquiring knowledge of spatial lay out of the maze and this knowledge is capable of revealing itself in a variety of different ways.
1.11.2. Learning varieties:
There is one major distinction between types of learning concerning the complexity of the information to be learned. The following list is a hierarchical order of the types, beginning with the simplest. Although this study concentrates on human learning, organisms possessing fairly well developed neural systems have proved capable of learning.
First one is "Relational" learning, the learning of a relation between two stimuli or between a stimulus and behavior required. Concerning this one there are many subtypes the simplest one is the classical conditioning which involves learning specific relations between environmental stimuli, and there are more complex types of relational learning. Nonassociative learning is obtained by repeated exposure to a single stimulus type; associative learning comes from comparison of one stimulus to another (classical conditioning), or comparison of a stimulus to the organism behavior (operant conditioning). Knowledge can also be acquired by repeated practice (reflexive learning).
Second is "Nonrelational" learning, only a single stimulus (an environmental event) is involved, and the simplest form of nonrelational learning is "Habituation", we have all experienced this type of learning when frequently occurring nonnoxious stimuli such as household noise decrease to attract our attention. And the second form of nonrelational learning is "Sensitization" in which an organism learns to increase the vigor of a response after exposure to a noxious or threatening stimulus. We can say it is reverse habituation but it is not exactly dishabituation.
Motor learning this is the learning of skilled motor tasks e.g. tying shoelaces. It arises through a combination of sensory ability, physical dexterity and cognition. A sequence of two more stimulus-response associations is ‘chained’ together resulting in the motor skill.
Language and verbal learning is concerned with the acquisition of human language, similar to sensorimotor learning in the sense that it too uses ‘chaining’, albeit on a verbal level. It is best seen when learning a foreign language e.g. end (English)
® finish® fin (French)Only a small part of information entering the consciousness is stored, concepts and ideas can be stored as words (verbal memory) or non-verbal codes.
Categorical learning: This is learning to categorize objects and events. Categorical learning process of experiencing various stimuli, recognizing that some of them share important common traits and then grouping them together so that we react to them in similar way. Our ability to identify an object is dependent on this skill. Abstraction is the process of recognizing that certain stimuli, which share similar attributes, belong in a particular category. Generalization allows us to recognize new instances of a category when encountered. The difference between two objects may the incredibly subtle yet an adult can classify more than 100.000 items.
For example, every instance of a fish encountered is stored as a featural description (exemplar) in memory a new object is analyzed by the sensory system to create a list of features that allows comparison with stored exemplars. The new object is categorized as a fish if it is more similar to the stored exemplars of fish than exemplars of others categories.
Three types of categories are used in categorization. Conjunctive categories have members identified by the presence of two or more memberships. In disjunctive categories membership is based on either the presence of one attribute OR the presence of another attribute. Relational categories do not simply use the presence or absence of attribute but rather a relationship between attributes.
Rule learning: this is the highest form of learning and as such recruits the most advanced perceptual skill.
Perceptual learning refers to such things as recognizing or distinguishing between objects or stimuli in the past from their visual appearance, how they feel, how they smell, or their sounds, each of our sensory systems capable to perceptual learning. We can also recognize surrounding peoples from their faces, movements, voices, and we can recognize their emotional status by recognizing the word they are saying. Learning visual stimuli recognition involves changes in visual association cortex, by receiving information from lateral geniculate nucleus of the thalamus to the primary visual cortex (Striate cortex). Within it individual’s modules of neural analyze information, V3 is devoted to analysis of orientation, area V4 to analysis of colors, and V5 to analysis of movements. Then these sub-regions of prestriate cortex send the results to the Inferotemporal Cortex, which combines the informations and produces neural activity that corresponds to the perception of particular three-dimensional stimuli. Auditory stimuli associated with changes in auditory cortex and so on. Perceptual learning seems to be accomplished by changes in the sensory associated cortex.
Reber, 1965, 1967, & 1969 introduced the term of implicit learning, which was about characterization of how one develops intuitive knowledge about the underlying structure of a complex stimulus environment. He argued that the characterization of the implicit learning is basically by these two critical features. First is that the implicit learning is an unconscious process, and the second is that it yields abstract knowledge, stimulus environment display and abstract representation of a structure induction lead to implicit knowledge.
Since then a lot of evidence to support the claim that sequence learning may occur without concurrent awareness of sequential structures has been based on dissociation between performance increments in the serial reaction time task and performance in measures of conscious knowledge. The first, reliable performance increments in serial reaction time tasks have reported subgroups of participants who were not able to verbalize the sequence structure after training or who had not even noticed that there was a structure (Cohen, et al. 1990; Curran & Keel, 1993; Reed & Johnson, 1994). Second, participants showed learning of serial reaction time tasks even if their performance in a subsequent prediction task in which they had to predict on each trial at which location the next stimulus would appear, was not superior to that of control participants who had been presented with a random sequence (Cohen, 1990; stadler, 1989; Willingham et al 1989). Third, sequence learning appears to be spared in amnesic patients suffering from Korsakoff’s and Alzheimer disease who show severe impairments of conscious episodic memory (Nissen & Bullemer, 1987; Nissen et al 1989).
There is some controversy about whether sequence learning is really unconscious. Perruchet et al, 1990; Perruchet, 1994; Shanks, & St. John, 1994. noted that the serial reaction time task was not well interpreted. Perruchet and Amorim 1992 conclude that the better way to assess by recognition and free-recall tasks for conscious knowledge about sequential structures.
1.11.3. Mechanism of learning:
Learning comprises three identifiably different dimensions: accumulation of knowledge, sequence of learning and varieties of learning.
The engram may exist as information in a reverberating circuit. (Dynamic engram), or as a modifications of synaptic connections (Structural engram).
Engram is the physical representation or location of a memory and known as memory trace. Hebb pointed out that memories can result from subtle alterations in synapses and these alterations can be widely distributed in the brain. Hebb postulated that learning and memory may rise from use-dependent alteration in the strength with which synapses transmit activity (Hebb, 1949). Synapses become strengthened as a result of persistent or repeated correlation between pre-synaptic and post-synaptic neuron activity.
Lomo 1966 first reported Use-dependent alteration in synaptic strengthen the rabbit hippocampus, eight years later, the term long term potentiation was used to characterize stimulus frequency dependent changes in population EPSPs and the amplitude latency of population spikes in the rabbit dentate, with and without anesthesia (Bliss and Lomo 1973). There may be changes in the proportions of nucleotides in RNA during learning, but attempts to extract and transfer these changes (as memory traces) between one organism and another have given inconsistent results and are now disregarded.
Accumulation of knowledge: Learning begins at birth when infant learns to associate feeding with the presence of parents. Children learn about their language. Adults attempt to use their accrued knowledge to comprehend the world and themselves.
1.11.4. Sequences of learning (3 stages):
Many researchers, especially the psychologists believe that learning sequences consists of three stages at least. The first one is Short Term Memory (STM), which roughly means storing a limited amount of information temporarily. The second stage is Long Term Memory (LTM), Which means storage of an information permanently, e.g. you could repeat seven dictated numbers, but you could not, in the same situation, repeat fifteen numbers. You could memorize fifteen numbers if you studied and rehearsed them for enough time, thus the simple way explaining how the entry of sensory information to the short term memory, then the conversion from short term memory to long term memory has been known as consolidation.
These stages are not necessarily independent of each other. In fact interaction between them may be essential. Past experience (stored in LTM) tells us what to expect and what to look for. Stimulus recognition is reliant upon information in LTM.
Practising simple visual tasks leads to a dramatic improvement in performing them. This learning is specific to the stimuli used for training. Ahissar, and Hochstein 1997, said that the degree of specificity depends on the difficulty of the training conditions. They found that the pattern of specificities maps onto the pattern of receptive field selectivities along the visual pathway. With easy conditions, learning generalizes across orientation and retinal position, matching the spatial generalization of higher visual areas. As task difficulty increases, learning becomes more specific. The dynamics of learning show a corresponding feature. Improvement begins with easy cases and only subsequently proceeds to harder cases. This learning cascade implies that easy conditions guide the learning of hard ones. The specificity and dynamics suggest those learning proceeds as a countercurrent along the cortical hierarchy. Improvement begins at higher generalizing levels, which, in turn, direct harder-condition learning to the subdomain of their lower-level inputs. They conclude that the learning can be effective using only difficult trials, but on condition that learning onset has previously been enabled. A single prolonged presentation suffices to initiate learning. We call this single-encounter enabling effect 'eureka'.
1.11.5. Where are learning does and memories themselves stored?
The answer seems to be that they are laid down in the cortex.
Karl Lashley 1920s conduct experiments to study the effects of brain lesions on the learning in rats. The studied question was how performance on this task was affected by making a lesion in same part of the rat’s cortex. Another experiment about the effect of the brain lesion location and size, He found in the first experiment that the rats given brain lesions before learning took more trials before they could perform the task. The second experiment concluded is that all cortical areas contribute equally in learning and memory.
Subsequent research has proven Lashley’s conclusion to be incorrect. All cortical areas do not contribute equally to memory. Lashley was correct that memories are distributed, and had an important and lasting impact on the study of learning and memory because he led other scientists to consider ways in which memories might be distributed among the many neurons of cerebral cortex.
Kluver and Bucy (1938) observed in animals that the complete removal of both temporal lobes resulted in a well-delineated syndrome known as the Kluver-Bucy Syndrome, which included a failure to recognize visual stimuli.
In 1954 Scoville described a grave loss of recent memory in human which he had observed as a sequel to bilateral medial temporal-lobe resection in one psychotic patient and one patient with intractable seizures. The removal extended posteriorly along the mesial surface of the temporal lobes for distance of approximately 8 cm. from tips of the temporal lobe and probably destroyed the anterior two third of the hippocampus and the hippocampal gyrus bilaterally. A human form of Kluver-Bussy syndrome is seen sometimes after herpes encephalitis, which damages the temporal lobe selectively.
The postoperative findings in 10 patients point to the importance of the hippocampal region for normal memory function (Scoville, 1957).
Lesions of the infero-temporal area are sufficient to produce visual memory deficit (Iwai and Mishkin 1968; Cowey and Gross, 1970)
Some evidence for this comes from pioneering experiments by Wilder Penfield (Penfield and Perot, 1963). But Penfield’s data have been criticized, for instance by Loftus (1980).
Modern brain researches have established that memories are encoded in the brain via the plasticity of synaptic connections between neurons, with individual memories being stored in parallel across huge numbers of neurons organized into "macrocolumn" in the cerebral cortex (Squire 1987). Each approximately 10 –11 neurons in the brain receive inputs from very many other neurons, and these inputs consists of neurotransmitter molecules that attach themselves on receptor sites on the dendrites of the neuron. When the input activation reaches a sufficient level, channels are opened which allow ions to be admitted into the neurons. These ions cause an electrical impulse to be generated, which then travels down the output pathway (the "axon") of the neuron, and which leads to neurotransmitter molecules being released, which can then act on other neurons.
When two connected neurons are excited at the same time, the synaptic connection between them may grow stronger, leading to facilitation between them. This is thought to be the basic way in which memory is stored. The process of synaptic plasticity has been extensively studied in the hippocampus, where so-called Long-term potentiation (LTP) has been observed. As a result of sending a train of impulses down a microelectrode attached to a part of hippocampus, increase in the strengths of connections are observed, such that a signal from one neuron elicits a much stronger response from another neuron than would normally be the case. These changes can last for a weeks or months. Although controversial, it is widely though the LTP is the basic process whereby the brain stores information.
Recognition memory may well depend on interactions involving the hippocampus, inferior temporal neocortex and the amaygdala, and retrieval perhaps more dependent on frontal-thalamic medial thalamic nuclei interaction (Joseph, R. 1996).
Frontal lobe: consists of (A) Primary motor cortex: Provides the main cortical output for the voluntary movement.
(B) Premotor cortex: Important for integration and programming of sequential movements.
(C) Prefrontal cortex: Covers a large area anterior to primary motor cortex and premotor cortex, divided into (1) Orbital frontal region (anterior tip of the frontal lobe), and (2) dorsolateral region. There are intricate connection between regions of prefrontal cortex and posterior cortex; the prefrontal cortex also has primary subcortical projections to and from the mediodorsal nucleus of the thalamus. (Goldman-Rakic, 1987; Fuster, 1989)
If there is prefrontal cortex dysfunction a variety of behavioral mental disturbances are seen in humans e.g. personality disorders, motor control & planning, language, problem solving, and memory impairment secondary to cognitive disorders such as deficits in attention, inferential reasoning, and cognitive mediation (Milner, et al 1985; Benton, 1991; Shimamura, 1991).
Three Occipito-temporal areas in the ventral object vision pathway had mostly transient responses to stimuli, indicating their predominant role in perceptual processing, whereas prefrontal areas demonstrated by sustained activity over memory. The posterior and inferior frontal gyri, the second inferior frontal gyrus and the anterior middle frontal gyrus (Courtney, et al.1997).
In a visual learning task, functional fields of activity appeared in the following anatomical structures:
The anterior cingulate cortex is assumed to play an important role in various aspects of human behavior, including affect, verbal expression, response selection, and initial action (Devinsky et al 1995).
The function of the cerebellum was traditionally described as motor, in the history of learning and memory, Thompson and colleagues brought attention to the role of cerebellum in classical conditioning and in the storage of memory engrams (McCormick & Thompson, 1984; Thompson & Donegan, 1986). More recently, neuropsychological, neuroimaging, and anatomical evidence has converged on the idea that the cerebellum also makes important contribution to cognition (Leiner, et al 1991. 1995). Cerebellar damage has been found to produce deficits in a variety in a cognitive measures, including intelligence (Botez et al., 1989), cognitive skill learning (Fiez & Petersen, 1993), in perception (Kolher et al., 1995), and working memory (Paulesu, et al 1993; Petrides et al., 1993).
There is a good evidence-in particular, from work on implicit learning in animals where Packard, and McGaugh (1996) concluded that:
McClelland & Goddard (1996) were discuss a framework for the organization of learning systems in the mammalian brain, in which the hippocampus and related areas form a memory system complementary to learning mechanisms in neocortex and other areas. The hippocampal system stores new episodes and "replays" them to the neocortical system interleaved with ongoing experience, allowing
generalization as cortical memories form. The data accounted for include first neurophysiological findings concerning representations in hippocampal areas. Second, behavioral evidence demonstrating a spatial role for hippocampus. Third, effects of surgical and pharmacological manipulations on neuronal firing in hippocampal regions in behaving animals.A memory separation, storage, and retrieval subsystem, supported by pathways between EC, dentate gyrus and area CA3, including the CA3 recurrent collaterals, which facilitates encoding and storage in CA3 of individual EC patterns, and retrieval of those CA3 encodes, in a manner that minimizes interference. A memory decoding subsystem, supported by the Shaffer collaterals from area CA1 to area CA3 and the bi-directional pathways between EC and CA3, which provides the means by which a retrieved CA3 coding of an EC pattern can reinstate that pattern on EC.
1.11.6. The principal brain areas involved in learning and memory (Fig 1.11):
The hippocampus that receives inputs from sensory cortex, and it is a part of circuit in which signals are transmitted from the subiculum (the "output" part of the hippocampus) to the mammillary bodies via a pathway called the fornix. From the mammillary bodies, signals are sent via the cingulate cortex back to the hippocampus. Although HM’s surgery removed structures in addition to the hippocampus, evidence suggests that highly localized lesions to the hippocampus, fornix, or mammillary bodies can create profound amnesia (Squire, 1992).
We can conclude the brain structure involved in memory when sensory inputs stimulate a cortical sensory area, cortex flow through parallel circuit to amygdala and hippocampus, both circuit encompass parts of diencephalon, prefrontal cortex; and dorsal forbrain, finally feedback to the sensory cortex closing the memory loop. The corpus striatum mediates the automatic connection between a stimulus and motor response.
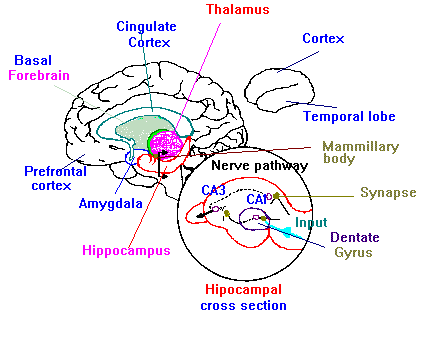
Figure 1.11 shows the principal brain areas involved in learning and memory.
1.12. MEMORY:
1.12.1. Types of MEMORY:
Working memory involves the short term maintenance of an active representation of information, and is responsible for the short-term storage and on-line manipulation of information necessary for higher cognitive functions, such as language, planning and problem-solving, and described as a buffer where a sensory inputs kept functional so that it can be used for further processing.
Implicit memory: is revealed when previous experiences facilitate performance on a task that does not require conscious or intentional activation of those experiences.
Explicit memory: is revealed when the performance on a task requires conscious recollection of previous experiences.
Sensory memory: Sensory information (Visual received by retinal receptors) stored automatically for a few 100msec are evaluated and forgetting by. Placement of other information immediately processed verbal or non-verbal or distinguished.
Primary memory: Short-term memory verbally coded information is temporarily stored in the order received, repetition ensures the transfer of the contents of the primary memory to the secondary memory. New information may arrive before transfer to secondary memory, hence the original information is forgotten
Secondary memory: Long term memory stored in order of significance as engram previous experience or information acquired subsequently proactive or retroactive informations may interfere with the learning processes. So partial or complete forgetting may occur proactive inhibition is important bin causing memory loss.
Tertiary memory: Important information is stored indefinitely, is rarely forgotten, rapidly retrieved (e.g. an individual name).
Two types of working memory are thought to exist. A brief mental after image lasting hundreds of milliseconds and a longer less literal vivid recollection of sensory features for up to thirty seconds. Evidence for this can be seen in Sperling’s (1960) ingenious experiment. Just after a complex image was displayed, auditory clues were given to indicate which part of the image to remember. If this clue was given within 250 msec of the image it aided memory, suggesting that aspects of working memory be converted to categorical within this time. Further evidence is shown by the fusion of TV pictures. Successive visual stimuli within 200 msec fuse perceptually (Haber and Standing, 1969 & 1970). Briefly presented stimuli are masked by a subsequent pattern presentation within 200 msec (Turvey, 1973). It has been hypothesized that trace develops with attention. After the construction of an automated tool of stimulus processing, that is automatic in the absence of attention (Näätänen and Alho, 1995).
Traditionally, working memory processes have been divided into two types, the first one is an executive control (governing the encoding manipulation and retrieval of information in working memory) and the second one is an active maintenance (keeping information available on line). It has also been proposed that these two types of processes may be subserved by distinct cortical structures, with the prefrontal cortex housing the executive control processes, and more posterior regions housing the content-specific buffers (for example verbal versus visuo-spatial) responsible for active maintenance. However studies in non-human primates suggest that dorso-lateral regions of the prefrontal cortex may also be involved in active maintenance. Working memory involves the short-term maintenance of an active representation of information so that it is available for further processing.
Visual working memory tasks, in which subject retain the memory of a stimulus over brief delays, require both the perceptual encoding of the stimulus and subsequent maintenance of its presentation after the stimulus is removed from view.
Visual memory tasks require both the perceptual encoding of the stimulus and a prolonged maintenance of its representation after removal from view. These two mechanisms are, an executive control responsible for encoding and retrieval of information in working memory, and an active representation consists of a visual sketchpad and phonological loop (Cohen et al, 1997).
The visuospatial sketchpad is assumed to be the comprised of separable visual and spatial components, serving different functions, and these functions is also consistent with recently proposed models of two mental imaginary, one that preserves the appearance of objects including color and form information, and second spatial component that preserves representation of layout (location & size) of objects in space.
Mismatch negativity (MMN) has been seen as a response to change in an ongoing series of tones and thus linked to working memory. MMN absence using inter-tone delays of more than two seconds, suggests that the representation was transient (Mantysalo and Naatanen, 1987). In recent study evidence for activation up to ten seconds has been found (Czigler et al, 1992). The similarity between these timing and behavioral discrimination tasks suggests that the internal representation is sensory.
During stimulus encoding certain stimuli can produce memory representation or codes of multiple types, e.g. printed word can yield separate lexical, orthographic and phonological codes. This ‘sensory’ or working memory cannot be equated with particular type of stimuli but rather the type of codes produced in the brain.
Such tasks activate multiple areas in visual and prefrontal cortices. To delineate the roles these areas play in perception and working memory maintenance, we used event related potentials ERPs to obtain potentials measures of neural activity related to different components of event working memory categorical task, non selective response to visual stimuli, selective transient response to images and sustained response over memory delay.
The systemic progression in relative strength of perception and memory related activity from the posterior extra-striate through to prefrontal areas would suggest that the neural system for working memory be hierarchically organized. The prefrontal regions also are activated during long-term memory recall. Active presentation of recalled material might mimic working memory images. The temporal cortex shows a similar sustained activation but is disrupted by further stimulus, unlike the frontal cortex (Miller et al, 1996). This implies that the temporal cortex is predominantly perceptual but helps in working memory unless recruited for perceptual tasks.
Experiments by Cowan (1984 & 1988) show working memory include a transient nature for comparison of two consecutive stimuli. Discrimination declines as inter-stimulus time increases up to 30 seconds. Although some long-term components are present e.g. the end performance is greater than the chance (Cowan, 1995).
The object property pathway runs from occipital lobe to inferior temporal lobe and spatial properties pathway runs from occipital lobe to parietal lobe and both of them called what system and where system respectively. There are some physiological evidences about increase the regional blood flow (rBF) to superior parietal cortex with spatial task (dot location matching), and with face matching task, the rBF was towards the posterior temporal cortex and lateral occipital area was involved in the tasks (Haxby et al. 1991).
Functional MRI Imaging studies have shown that both visual and prefrontal cortices are active during visual memory task. Work using these techniques with face discrimination task, has shown that the ventral Occipito-temporal extra-striate visual areas are active during stimulus representation. A region in the posterior lingual and fusiform gyri showed a non-selective response to stimuli. The anterior fusiform gyrus activity was face stimuli dependent, and suggested that it was concerned with perceptual processing of faces and could hold the active representation. Three prefrontal areas have been connected with working memory, The posterior middle and inferior frontal gyri, The second inferior frontal gyrus and the anterior middle frontal gyrus, showing sustained activity during memory delay (Courtney et al, 1997).
1.12.2. Memory disorders
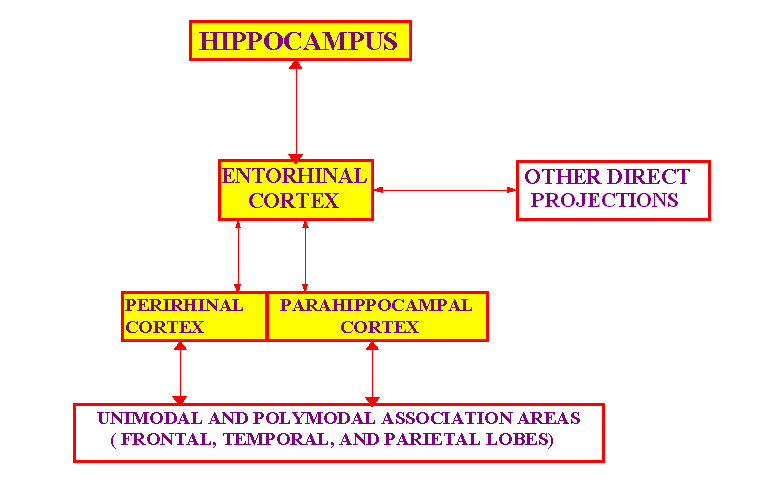
Figure 1.12 shows a schematic view of the brain structures, and the important connections for the declarative memory. Yellow areas indicate the structure within the medial temporal lobe
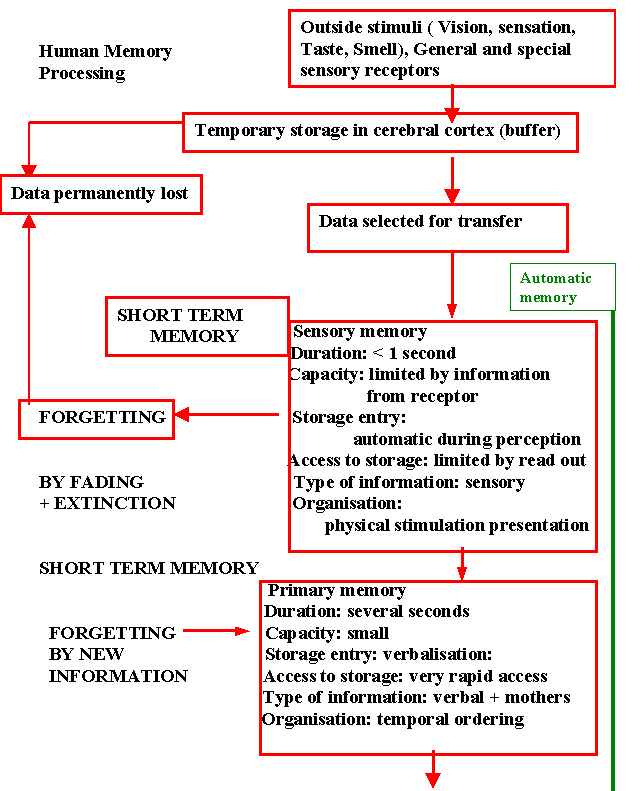
Figure 1.13a shows the human memory processes (short-term memory)
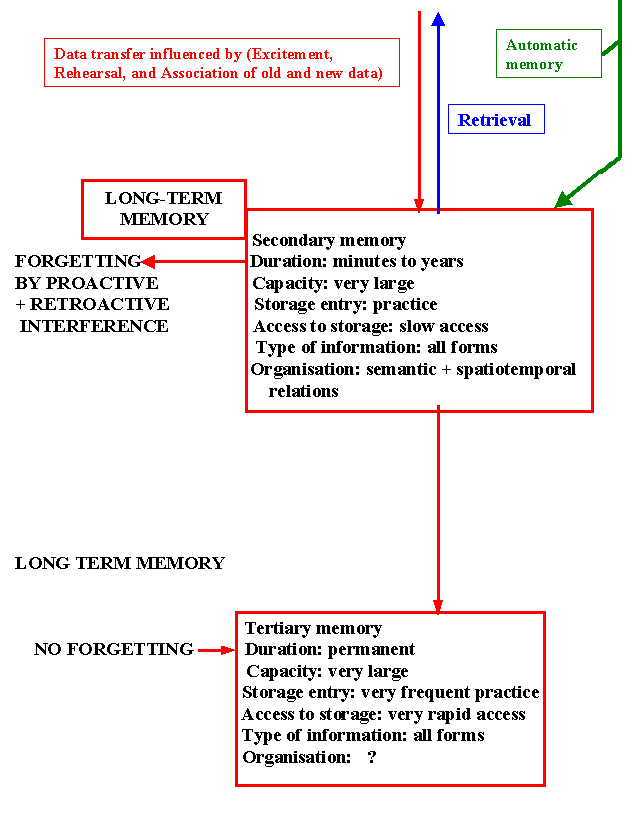
Figure (1.13b) shows the human memory processes (long term memory)
1.13. Categorical Perception:
1.13.1. Categorical perception:
Since the beginning of the experimental study of categorisation in psychology (Hull 1920), there has been a tendency to assume that all acts of categorisation are accomplished by the same means.
Who can imagine a world without categories? Categorization helps to reduce the complexity of the environment, identify objects, reduce the need for learning, relate objects to a hierarchy of knowledge, and help make decisions.
Categorical perception occurs when the continuous, variable, and confusable stimulation that reaches the sense organs is sorted out by the mind into discrete, distinct categories who members somehow come to resemble one another more than they resemble member of other categories.
In general various investigators have defined the categorical perception in terms of identification performance, discrimination performance, or both. Simply there is no uniform definition of categorical perception (Medin, D.L. and Barsalou, L.W. 1987).
Categorical perception (CP) is a process that allows the brain to place continuous variables that are transuded by the sense organs into discrete, distinct, categories. Categories are members somehow come to resemble another one, more than they resemble members of another different categories, or by another words members within groups resemble each other more than across boundaries. The perceptual difference inter-category can be more than suggested by the actual physical attributes. Consequently the perceptual similarities within a group are increased. A well used example of categorical perception is that the color continuum.
Physically the spectrum of light that is perceived as color varies in a linear way. Although the light wavelength varies in an analogue fashion, we classify the colors as distinct groups with boundaries. This means that a red and orange can be physically closer in wavelength than two oranges but are perceived as different categories. Unlike categorical perception of color, which is not present at birth, and unill six weeks after birth due to late demylination of the system. Infants learn to tell the difference between cats and dogs using the same grouping technique. A child of three may see a lot of similarities between the two classes and struggle to qualify individual examples (Harnad, 1987)
Chicken sexing has been quoted as a working example. A difficult task that has to be learnt so that two groups can be reliably distinguished, categorical perception is thought to be a basis for cognition and thus learning.
The phenomenon of categorical perception was first suggested within each perception research (Liberman et al, 1957; Liberman et al, 1967), unto which it was originally thought unique. Although now CP. has been proved to exist in other contexts. CP can be seen as an analogue to digital conversion. Where qualitative differences and perceptual boundaries have arisen between groups.
CP and continuous perception were initially thought of as two distinct processes, even isolated to left or right hemisphere (Liberman et al, 1967). Psychological models have been proposed that expand on this distinction. Braid (1969) defined a model with two parameters, trace and context. Trace referring to the comparison of the stimulus to a previous sensory working memory image. The context to certain anchor features. From this model you can see that short term, long term and working memory are involved.
Harnad (1987) has asked two questions considered as the most basic questions about the human or the animal perception and cognition. The first one is "how do we sort objects, people, and ideas in the world into their proper categories? The second is "What transforms the "booming, buzzing confusion’ that enters our eyes and ears at birth into that orderly world we ultimately experience and interact with?
Perception refers to the means by which information acquired from the environment via the sense organs is transformed into experiences of objects, events, sounds, tastes, etc.
Perception is a very complex process, depends on physiological systems associated with each sense modality, plus central brain processes, which integrate and interpret the output from the physiological systems (Frisby. 1979).
1.13.2. Theories of Category classifications:
Feature Comparison theory of category classification: A category (or concept) is defined by matching the features of an object to a logical arrangement of features in a category (stored in our memories). All features are considered equally good and necessary. All category members are equally representative.
Types of feature rules used in categorizing:
Prototype theory of category classification: Theory Basics: It is suggested that categorization is based on similarity of features in perception and memory, but that none of the features are necessary for defining. Generally speaking, the prototype of a category contains characteristics attributes of its category exemplars, namely, attribute that highly probable across category members, but that neither necessary or sufficient for category membership.
Exemplar theory of category classification: People classify entities on the basis of their similarity to memories of previously experienced category members. They compare it with memories of specific category exemplars, each memory representing an encounter with an exemplar at specific place and time. Stimuli are assigned to the category having the most similar exemplar or exemplars.
Boundary theory of category classification: A final classification strategy is that people determine category membership on the basis of category boundaries rather than characteristics or ideal attribute.
1.13.3. Where is Categorical Perception executed?
There is no one area where we can look for CP. activity. We are concerned with a system that allows two way interactions between the current stimulus and ongoing evaluation of perception. The new stimulus has to be incorporated into the perceptual model, but also has to be perceived in the cortex of this model. That is affective and effective process occurring within the same mechanism.
In primate there are cells within the occipital striate cortex that respond faithfully to analysis of physical dimensions such as length, width and orientation (Hubel and Wiesel, 1968). Areas of a visual stimulus within the prestriate cortex contain multiple representations of the visual fields for further analysis (Zeki, 1978). Visually responsive areas exist in the inferior portions of the temporal lobe (Gross et al, 1972).
To add another dimension to the processing that possible, associative cortical areas receive a secondary neural input from the thalamus as well as the visual sensory area (Diamond, 1979). This enables a two way system as suggested involving parallel projections from clusters of thalamic nuclei that serve different perceptual functions (Benevento and Rezak, 1976). For example the multiple connections of lateral and inferior portions of the pulvinar nucleus of thalamus are afferent and efferent to the visual cortex. Thus allowing reverberating sustained circuits.
Categorization plays a critical role in perception, thinking, and language and is probably a significant factor in motor performance.
1.13.4. Effect of brain damage on Categorical Perception:
In animal studies monkey inferotemporal lesions lead to loss in categorization. Complementary to the striate lesions which cause a loss in acuity not CP (Wilson and DeBauche, 1981).
In human examples, CP of visuospatial and tactuospatial continua were more affected by damage in the right hemisphere (Bertelson, 1982). There are many examples of the right side of the brain playing an important and independent part in categorical perception.
1.14. Cognition
The word cognitive comes from the Latin word cognoscere, meaning to know and this indicates what cognitive psychophysiology is all about.
Cognitive psychophysiology tries to explain how the human mind comes to know things about the world around it people, and about itself, and how it uses this knowledge to perform an impressive range of tasks such as remembering, speaking, performing skilled action, solving problems and reasoning.
Cognitive science is the study of the human mind as an information processing system. As such it seeks to utilize any method in its power to quantitatively model the mind. It seems that a huge aspect of what it is to be human is ignored by this definition, as humans are both spiritual and emotional beings. From my point of view cognitive science deals only with one of several aspects of what it is to be human. To take its concepts and beliefs, they are an absolute model of how people are, seems to me to be very dangerous, as this would lead to a very mechanistic view of people which manifestly we are not.
1.15. Attention:
There is one thing for sure about attention. It is not something concrete inside our body, it is not directly observable, and it is a cognitive process. Attention is a hypothetical construct, which accounts for the selective capacity of humans and animals to respond to certain stimuli and ignore others that appear equally potent on the physical and the time dimension.
Attention is the taking possession by the mind, in clear and vivid form of one out of several simultaneously possible objects or trains of thought. Perceiving, and remembering need attention and attention is initiated by appraisal that this is good to know, good to do, and want to do-good to want result in wanting and approaching it. One of the most common daily activities is to select something to focus on. That is the operation of attention! The basic assumption is that we are limited in only being able to focus on a few things or carry out a few skills at the same time. Attention is switchable from one focus to another. Two types of attention models, namely the filter (or bottleneck) models and the capacity model.
The fact that we recognize structures in our environment without any effort, although these structures are always embedded variety of visual contexts, actually we use two different attentional systems voluntary or active (endogenous), which involves the attention-paying process that results from intention, instructions, etc. Involuntary or passive (exogenous) which involves the attention getting process that results from the intrinsic properties of a stimulus and usually defined in terms of orientation responses.
1.16. The Mind
The word mind is frequently use as noun in English. If the mind is a noun we tend think that it must refer to a thing, but it is a strange one if it is a thing. It has no weight, color, extension, and it is non-physical. How can such a thing exists and how can it affects and be affected by the brain? It is an abstract noun.
But if we consider that thing as a set of processes and activities it will be easier. Such a word as "respiration" refers not to single thing but to physiological biochemical processes and activities starting with inspiration...expiration. These steps or processes and activities can take place because there exist physiological organs, such as mouth... trachea...etc., which support them and make for interaction of biochemical processes.
In similar way we can think of there being a set of mental processes carried out by mental organs, the brain, which are needed to carry out activities such as memory, language, and thinking.
Mind is better described as a set of processes, which rely on the brain, rather than as a separate, mysterious thing. These processes relate to what a person says or do, and are linked to observable behavior, which allows us to test theories about inner mental processes by observing their effect on outward measurable behavior
To have a mind is to have feelings. Not just feelings like pain or anger etc., feelings like what it feels like to be hungry, to see a red chair, to want to eat, to imagine a pink elephant and what it feels like to believe that the earth is round, a sentence understanding, a word meaning, etc.
In short, to have a mind is to be capable of having experience (because there's something that it feels, like to experience each of the many things we are able to experience). Things like rocks and computers have no minds: they do not have any experiences.
There is no exact definition of the mind - it is a very personal concept. Some people may believe that the mind is conscious awareness, or even a soul. It is a consolidation of all the different mental process that is occurring at one time - e.g. vision, hearing, speaking. Another definition of 'the mind' may be having a theory of mind - being able to understand other's thoughts and feelings.
Who can prove that we have mind. Only that we ourselves have a mind, but we can never know that someone else has a mind, surely all of what we are studying can said to be brain related!
This lead to the well-known Descartes famous conclusion:
"Cogito ergo sum"..."I think therefore I am"